ПОСТПІРОГЕННІ ДЕМУТАЦІЇ ЕКОСИСТЕМ НАДРІЧКОВИХ ПІСКІВ НА МИКОЛАЇВЩИНІ
Июн 16th, 2009 by stepan
УДК 502.7:581.9:631.95 (477)
ПОСТПІРОГЕННІ ДЕМУТАЦІЇ ЕКОСИСТЕМ НАДРІЧКОВИХ ПІСКІВ НА МИКОЛАЇВЩИНІ
Г.В. КОЛОМІЄЦЬ, аспірант,
Р.І. БУРДА, доктор біологічних наук, професор, Національний аграрний університет, ННІ охорони природи і біотехнологій
Досліджено постпірогенні демутації свіжих згарищ на надрічкових пісках в нижній течії Південного Бугу та Інгулу, а також Кінбурнського півострову. Виявлені видове багатство, біоморфологічна та ценотична різноманітність фітобіоти пірогенних екосистем на місці штучних насаджень Pinus sylvestris L. та P. pallasiana D. Don. на колишніх псамофітних степах. З’ясовано: видове багатство псамофітону у певних межах відновлюється протягом 3 – 7 років; демутація відбувається за участі видів, які збереглися у соснових культурах, та міграцій з навколишніх степових ділянок; фітосозологічна цінність екосистем посилюється (13 видів, що підлягають особливій охороні), проте, участь випадкових видів, бур’янів становить 44 %, а серед них 73 % – адвентивні. За диференційованого підходу згарища цілком придатні для використання в розбудові локальних ланок екомережі Миколаївської області в категорії «території відновлення (ренатуралізації)», а з часом окремі з них – «території природного розвитку».
Постпірогенні демутації, згарища, псамофітон, видове багатство.
Постпірогенні сукцесії екосистем вивчені у різних біомах досить детально. Пожежі вважаються провідним екологічним чинником в еволюції низки екосистем [7, 13]. Формування біоти багатьох з них відбувається під контролем природних пожеж, свідченням чого є деякі адаптації, наприклад, рослини-пірофіти [2]. Поряд з природними пожежами з прадавніх часів вогонь є антропогенним чинником (спрямовані пали при масових полюваннях, підсічно-вогневе землеробство, весняне випалювання лучних і степових пасовищ і сіножатей тощо). Вплив антропогенних пожеж уже спричинився до виникнення мільйонів гектарів розріджених чагарникових пустищ у Середземномор’ї, знеліснення з наступним опустеленням у Африці. Нині підсічно-вогнева система землеробства є загрозою для тропічних лісів на усіх континентах [6]. Підкреслюється високий елімінуючий вплив вогню на біоту. Через раптовість і короткотерміновість пожежі, зазвичай, не призводять до повного вимирання усієї біоти. Відновлення біоценозу відбувається за рахунок стійких до вогню видів, рослин-пірофітів, з підземними бруньками відновлення (геофітів), збереження насінням схожості на згарищах, рясного обсіменіння, здатності рослин-експлерентів освоювати свіжі згарища, активного або пасивного розселення тварин, які знаходять сприятливі умови для життя на свіжих демутаційних згарищах тощо.
Вогонь діє спряжено на абіотичні і біотичні компоненти екосистеми, викликаючи ланцюги послідовних дигресивно-демутаційних змін. Це є підставою для виділення постпірогенних сукцесій екосистем. Ю. Одум [13] зміни екосистем після пожежі називає «пульсуючими стабільними сукцесіями». Більше того, Т. Работнов [14] вважає, що там, де вогонь є чинником еволюції, випалювання не можна виключати з сукцесійного ряду аж до екосистем з режимом заповідання. Знавець сукцесій в українських степах В. Ткаченко [15] рекомендує включати контрольовані пали до регуляційних заходів у степових природних заповідниках з метою відтворення еталонних якостей степів. У згадуваній уже праці А. Тишков [7] посилається на практику заповідної справи США, де ще у 1975 р. було прийнято програму «направлених локальних випалювань» (пожежі догляду – «management-fires» і дикі пожежі «wild-fires»). Спрямоване випалювання заростей карібської сосни в національних парках Еверглейдсе (Флорида) розпочато ще з 1953 р.
Підсумуємо цей короткий огляд. Біоті більшості біомів певною мірою притаманні адаптації до вогневого впливу, а постпірогенна демутація відбувається швидко. Цьому сприяє достатнє забезпечення продуцентів елементами мінерального живлення в перші роки після пожежі та підвищення на згарищах якості пасовищ для фітотрофів.
Дослідники надрічкових і приморських пісків України, вивчаючи процеси заростання голих пісків, розглядають демутаційні процеси як дуже складні і тривалі [3, 10]. Заміна одних угруповань іншими відбувається повільно і малопомітно, як наслідок – виділення стадій заростання доводиться проводити умовно. І. Гордієнко [3] подав процес заростання Олешських пісків у комплексі факторів: життєдіяльність ентомофауни і хребетних тварин, діяльність вітру, ценотичні умови тощо.
На тлі наведених поглядів провідних екологів, в цій статті зроблено спробу аналізу відновлення фітобіоти при демутації екосистем на три-семирічних свіжих згарищах у Миколаївській області та з’ясування придатності їх для розбудови регіональної ланки національної екомережі. В контексті принципу суцільної охорони екосистем ландшафтного рівня, як це передбачає Європейська конвенція про ландшафти [5], вирішення цих завдань є актуальним в екології і раціональному природокористуванні.
Місце, обсяг і методика досліджень. Місцем для вивчення постпірогенних демутацій обрано надрічкові піски в нижній течії Південного Бугу, Інгулу та Кінбурнський півострів. За певного комплексу природних та антропогенних чинників, зокрема, надмірного пасовищного навантаження наприкінці ХІХ ст. на Кінбурні утворились рухливі піски. Для закріплення їх на засадах біогеоценотичного підходу запроваджено низку радикальних заходів [3, 10]. Серед них найважливішими було визнано створення штучних деревних насаджень, у тому числі сосен звичайної (Pinus sylvestris L.) та кримської (P. pallasiana D. Don.). Наразі на Нижньодніпровських пісках майже 100 тис. га штучно створених соснових насаджень, з яких у Миколаївській області 5 тис. га (Кінбурнський півострів). Вони мають невисоку продуктивність, уражаються шкідниками, пожежно небезпечні, подекуди всихають у молодому віці. Державна комісія в 1998 р. прийняла рішення про скорочення площ штучних соснових насаджень в Нижньому Придніпров’ї. У 2001 – 2002 рр. пожежею було знищено понад 1 тис. га лісових культур Кінбурнського півострова. У другій половині ХХ сторіччя соснові насадження були створені на пісках у пониззях Південного Бугу та Інгулу; вони так само страждають від пожеж. На підставі актів обстеження згарищ у місцезростаннях вузькоендемічних видів та згарищ штучних насаджень сосни у межах природно-заповідного фонду, що складені за нашою участю О.М. Деркачем, С.В. Таращуком, М.М. Парафіло, а також додатково проведених нами у 2004 – 2007 рр. досліджень описано фітобіоту 51 згарища (табл. 1). Обліки проводились шляхом послідовної фіксації наявності-відсутності виду у межах локалітету. У кожному локалітеті закладались 30 облікових ділянок розміром 10 х 10 м. Для семи созологічно цінних видів, крім того, підрахована чисельність особин на 1 м2 у межах трьох об’єктів природно-заповідного фонду.
Трапляння установлено як частка (%) облікових ділянок, на яких зафіксовано наявність виду, до загального числа облікових ділянок у певному локалітеті. Наукові назви видів приведені відповідно до вимог чинного Кодексу ботанічної номенклатури [17]. Усі таксономічні і типологічні аналізи проведено з використанням власноруч створеної бази даних «Екомережа» [8].
1. Перелік згарищ у Миколаївській області, на яких вивчено постпірогенні демутації фітобіоти
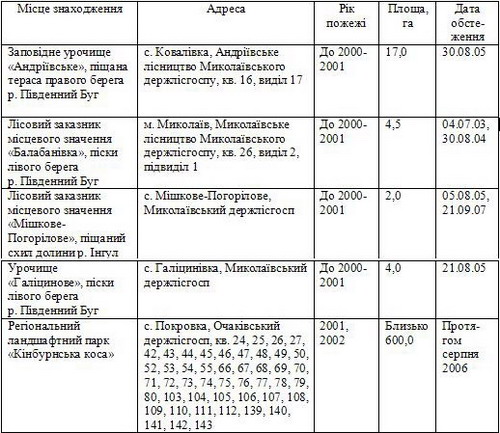
Результати та їх обговорення. Фітобіоту постпірогенних екосистем складають 359 видів судинних рослин з 217 родів та 50 родин. Середнє видове багатство родини – 7,2 – менше, ніж в середньому для Миколаївської області (9,3), середня чисельність видів в одному роді – 1,7, в області – 2,3, що пояснюється не лише малою площею згарищ, але й меншою різноманітністю екотопів в їх межах.
На підставі аналізу типологічної різноманітності фітобіоти за класифікацією життєвих форм Раункієра встановлено наступне. Серед 359 видів вивчених згарищ переважають види, бруньки відновлення яких за несприятливих умов середовища перебувають під ґрунтовим покривом (геофіти – 10 %) або розташовані у великій кількості на поверхні ґрунту (гемікриптофіти та хамефіти – понад 40 %) і здатні відновлюватись вегетативно. Зокрема, вже на 12-й день після пожежі 2001 р. на Кінбурні було зафіксовано відростання від підземних бруньок Betula borysthenica Klokov, Populus tremula L., Salix acutifolia Willd., S. rosmarinifolia L. Ушкоджені вогнем дерева Pinus sylvestris та P. pallasiana гинуть, природне поновлення їх за умов згарищ не відбувається, проте в літературі види роду Pinus L. відносять до пожежестійких [7, 13]. З цього випливає, що понад половини видів, наявних в пірогенних екосистемах, імовірно, відновились за рахунок особин або діаспор, що збереглися після пожежі. Решта, особливо терофіти (41 %), імовірно, є мігрантами, діаспори яких занесені з прилеглих залишків псамофітних степових угруповань.
Проте, не слід сподіватися, що за такий короткий термін і на таких зовсім незначних площах своєрідний флороценотип, яким є псамофітон надрічкових пісків, відновився повністю. У складеному нами переліку лише 244 види (68 %) є аборигенними. Третина видів – адвентивні, в т.ч. – культигени, а серед аборигенних 42 є випадковими – вони рудеранти, які разом з адвентивними формують новітній флороценотип – синантропофітон [1]. Переважання видів степо- та псамофітону свідчить про високий потенціал пірогенних екосистем для природного відновлення псамофітних степів. Ця теза правомірна стосовно як фонових, так і рідкісних, ендемічних видів. Більше того, трапляння та рясність видів, що підлягають особливій охороні на загальнодержавному та світовому рівнях, вражаюче високі (табл. 2, 3). У штучних соснових насадженнях, під пологом сосни за нашими спостереженнями ці види не здатні підтримувати свої популяції [4].
2. Види рослин, що підлягають особливій охороні, у фітобіоті пірогенних екосистем*
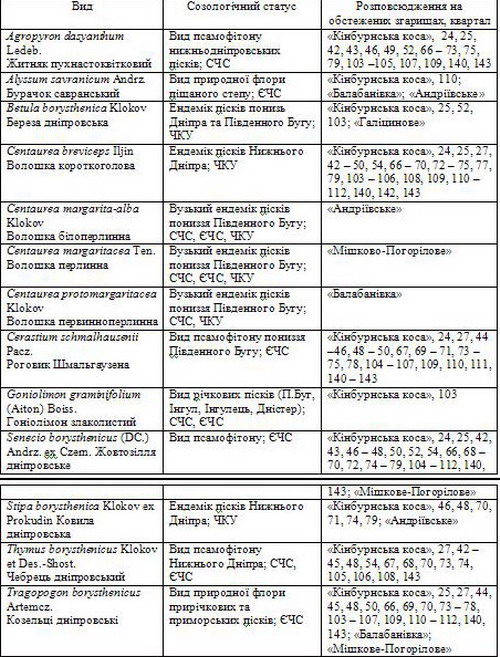
*Примітка. У колонці 2 позначено: СЧС – вид занесений до Червоного списку IUCN [12], ЄЧС – вид занесений до Європейського Червоного списку [16], ЧКУ – вид занесений до Червоної книги України [16].
3. Трапляння та рясність деяких видів фітобіоти пірогенних екосистем
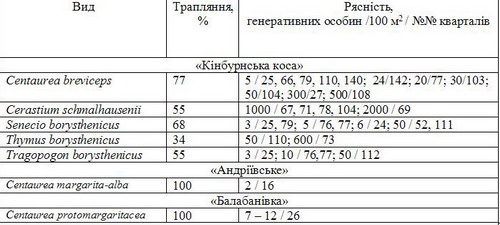
Крім видів, що підлягають особливій охороні, при обстеженні влітку 2006 р. в усіх кварталах виявлені фонові види піщаного степу пониззя Дніпра – Artemisia marschalliana Spreng., Achillea micrantha Willd., Carex colchica J. Gay, Helichrysum corymbiforme Opperman ex Katina, Festuca beckeri (Hack.) Trautv., Koeleria sabuletorum (Domin) Klokov, Polygonum arenarium Waldst. et Kit., Calamagrostis epigeios (L.) Roth, Dianthus platyodon Klokov, Jurinea laxa Fisch. ex Iljin, Linaria dulcis Klokov, L. genistifolia (L.) Mill., Jasione montana L., Scirpiоdes holoschoenus (L.) Sojak, Scabiosa ucranica L.
Такі високі темпи демутації обумовлені відновленням місцевих діаспор та проникненням їх з розташованих неподалік ділянок псамофітного степу. При цьому у соснових насадженнях, що існували тут до пожежі, залишки популяцій аборигенних видів присутні лише на галявинах [9]. Ці популяції не спроможні зберігати і відтворювати генофонд унікальних рослин псамофітону через невеликі площі. Пірогенні демутації стали важливим етапом відновлення природної фітобіоти, посилення созологічної цінності території. Зазначене, як приклад, підтверджується характером поширення на згарищах перлистих волошок (Centaurea margarita-alba Klokov, C. margaritacea Ten., C. protomargaritacea Klokov) (табл. 2, 3) – вузьких ендеміків пониззя Південного Бугу та Інгулу [11].]
Висновок
Спрямовані пожежі вважаються одним із важливих багаторазово випробуваних практикою форм регуляційних заходів, аж до екосистем, що підлягають особливій охороні. Ця роль пірогенного чинника стосується біомів різних природних зон. Керовані випалювання підтримують рослинний покрив первинних степів, які деградують в режимі абсолютної охорони, деяких типів хвойних лісів (сосняків, модринників), чапаралю, верещатників тощо.
З’ясовано, що протягом 3-7 років на згарищах спонтанно відновлюються популяції видів рослин, що властиві псамофітним степам із невеликими листяними гайками по зниженнях, які існували в цих ландшафтах до антропогенного втручання. Штучні насадження сосен не здатні до самовідновлення.
Висока рясність у пірогенних екосистемах фонових видів псамофітону як і видів, які підлягають охороні на загальнодержавному та світовому рівнях, підкреслює важливість пірогенних демутацій для відновлення природних екосистем псамофітних степів. Збереження їх оригінальності і генофонду є одним з найважливіших завдань Приморсько-степового (Азово-Чорноморського прибережного) екокоридору міжнародного значення. Викладене є незаперечною підставою для упередження щодо створення штучних лісових насаджень на згарищах без попередньої оцінки місцезростань і фітобіоти.
Список літератури
1. Бурда Р.И. Антропогенная трансформация флоры – Киев : Наук. думка, 1991. — 168 с.
2. Вальтер Г. Растительность Земного шара. – М.: Прогресс, 1974. – Т.2. Леса умеренной зоны. – 423 с.
3. Гордиенко И.И. Олешские пески и биогеоценотические связи в процессе их заростания. – К.: Наук.думка, 1969. – 242 с.
4. Збереження біорізноманіття в Приморсько-степовому екокоридорі. Каталог видів флори і фауни, що знаходяться під особливою охороною в Україні та є вразливими при здійсненні лісогосподарських заходів / Відп. ред. Таращук С.В. – К.: Громадська організація «Веселий Дельфін», 2007. – 112 с.
5. Європейська ландшафтна конвенція: Оф. перекл. – http:www.menr.gov.ua.
6. Екологічний потенціал наземних екосистем / За ред. М.А. Голубця. – Львів: Поллі, 2003. – 180 с.
7. Исаков Ю.А., Казанская Н.С., Тишков А.А. Зональные закономерности динамики экосистем. – М.: Наука, 1986. – 149 с.
8. Коломієць Г.В., Коломієць О.В. Приклад розробки географічної інформаційної системи для узагальнення досліджень заповідних територій та проектування екологічної мережі Миколаївської області // Роль природно-заповідних територій у підтриманні біорізноманіття : Матеріали наукової конференції, присвяч. 80-річчю Канівського природного заповідника (Канів, 9-11 вересня 2003 р.). — Канів, 2003. — С. 339–340.
9. Коломієць Г.В. Мережа природоохоронних територій Миколаївської області в контексті збереження фітобіоти // Агроекологічний журн., 2004. – № 3. – С. 13–18.
10. Косець М.І., Ткаченко В.С. Рослинність пісків // Рослинність УРСР: степи, кам’янисті відслонення, піски. – К.: Наук. думка, 1973. – С. 404–427.
11. Крицька Л.І., Деркач О.М. Сучасний стан популяцій видів ряду Margaritaceae Klok. (Centaurea L.)// Укр. ботан. журн. – 1991. – Т. 48, № 3. – С. 78 – 80.
12. Мосякін С.Л. Рослини України у Світовому червоному списку // Укр.ботан.журн. – 1999. – 56, № 1. – С.79–88.
13. Одум Ю. Основи екології. – М.: Мир, 1975. – 740 с.
14. Работнов Т.А. Фитоценология. – М.: Изд-во МГУ, 1978. – 296 с.
15. Ткаченко В.С. Фітоценотичний моніторинг резерватних сукцесій в Українському степовому природному заповіднику. – К.: Фітосоціоцентр, 2004. – 184 с.
16. Червона книга України. Рослинний світ / Редкол.: Ю.Р. Шеляг-Сосонко (відп.ред.) та ін. – К.: «Українська енциклопедія» ім. М.П. Бажана, 1996. – 608 с.
17. Mosyakin S., Fedoronchuk M. Vascular plants of Ukraine. A nomenclatural checklist – Kiev: M.G. Kholodny Institute of Botany, 1999. – 345 p.


